编者荐语:
近年来,随着双抗药物研究的加速,各大公司纷纷布局,上期综述了双抗的挑战和开发策略,本期和大家从结构与功能优化方面谈谈双抗~更多精彩文章请见文末往期回顾。
多特异性抗体可以设计成多种不同的形式。二十多年来在双特异性抗体工程领域的研究和开发表明,抗体的设计和形式的选择对抗体的功能有着深远的影响。这尤其适用于引起细胞间生物学过程的反应,如受体激活、受体内化、受体聚集或在两个细胞之间形成免疫突触。为什么一些具有相同大小和结构域的形式比其他形式更有效?几何结构的重要性不言而喻,TriFab是一种新设计的抗体形式,由两个肿瘤细胞靶向实体和一个T细胞结合部分组成,特点是其紧凑的形状。TriFab与具有相同结构域序列的IgG样分子相比,这种独特的桶状形状使效价提高了35倍。
随着重组蛋白表达技术和抗体工程技术的发展,产生了许多不同的抗体形式。佳的抗体结构取决于所期望的生物效应及其潜在的结构条件。多特异性抗体用于各种目的,包括(1)受体激活(2)阻断(3)内化(4)聚集,(5)膜相关蛋白的结合,或(6)细胞毒性效应细胞的定向靶点(图1)。在这些应用领域中,下面几个例子证明了抗体形式对性能的影响。
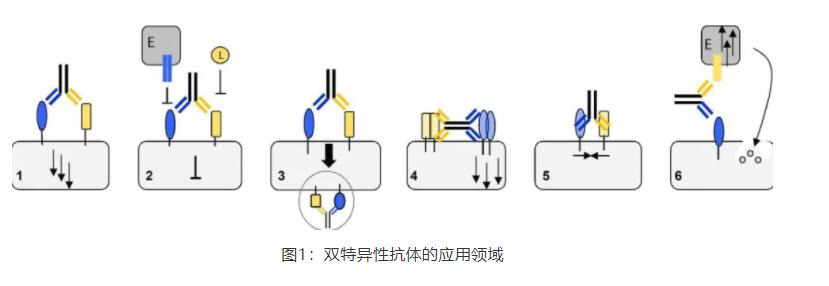
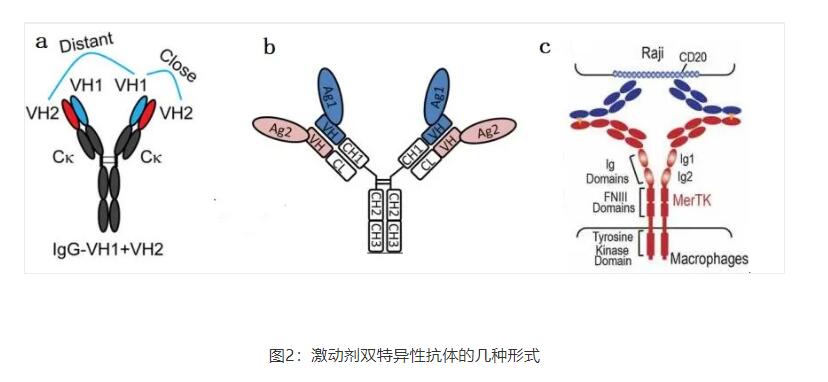
受体激活并诱导下游信号转导以达到一定的表型是激动剂抗体的目标。Shi等人提出一种IgG形状,四价,双翼形式,在每个N-端只包含VH结构域(图2a)。近距离的抗体结合部位模拟了天然配体,从而激活内分泌成纤维细胞生长因子FGF21受体(FGFR)。这种独特的抗体几何结构是经过精心设计的,并被证明是激活受体的关键。近也报道有同样形式的靶向CD40的激动剂抗体(图2b)以及一种二价的Y形Surrobody,激活死亡受体DR4和DR5。这种分子显示出比单一特异性DR4和DR5抗体更强的效力,这表明它具有独特的激活机制。近,Genentech的抗体工程师设计了一种简单的1+1IgG形式的分子(图2c),通过MerTK(Mer酪氨酸激酶)受体途径激活巨噬细胞,导致CD20阳性B细胞吞噬。
受体阻滞剂是目前肿瘤免疫治疗方法的重要组成部分。以抑制性免疫检查点为靶点的双特异性拮抗分子在克服肿瘤逃避机制方面显示出了很好的效果。一个突出的例子是免疫检查点的双重阻断,如LAG-3和PD-1,它诱导抗肿瘤免疫。正在进行的多种双特异性抗体形式的临床试验数量惊人。然而,相对于激动剂抗体的挑战性(它们依赖于亲和力和内在功效),设计阻断抗体主要依赖于寻找与天然配体竞争的高亲和力结合物,因此被认为并不太复杂。
抗体诱导的受体内化在抗体偶联药物(ADC)重定位中起重要作用。可以证明,与单价形式相比,EGFR的二价结合和由此引发的二聚作用导致受体-抗体复合物的内化显著提高。Niewehner等人分析了实现血脑屏障跨细胞作用的不同形式。转铁蛋白受体(TfR)的一价靶向导致体内脑暴露比二价形式高55倍。据推测,二价TfR结合导致大量受体二聚化,并随后进入溶酶体途径。另一方面,单价结合可使转铁蛋白同时络合,并将两种分子共转运至管腔的另一侧,而不会使抗体降解。
受体聚集的一个例子是激活由肿瘤坏死因子受体超家族(TNFRSF)的受体介导的外源性凋亡途径,例如死亡受体(DR)。诱导信号级联反应的激动性抗体的产生非常具有挑战性,在早期的方法中获得的激动剂抗体临床疗效有限。Yang等人用四价双翼抗体实现了稳定固有的激活,而二价双翼的对照分子没有触发任何激活。Brunker等人设计了以肿瘤抗原FAP(成纤维细胞激活蛋白)和DR5为靶点的2+2形式。结果表明,这种形式在顺式方向上的二价结合导致DR5亲和力驱动的高聚集,并随后表现出强诱导细胞凋亡的作用。与其他依赖FcγR相互作用的聚集方法不同,他们利用肿瘤间质中FAP的表达,有针对性地促进DR5的高聚集,从而降低FcγR依赖方法中的全身毒性。
另一个应用领域是抗体介导的磷脂膜上两种或两种以上蛋白质的结合。血友病A是一种由于凝血蛋白因子VIII缺失而引起的遗传性出血性疾病。在生理条件下,FVIII作为辅助因子促进酶-底物复合物FIXa-FX的结合,从而激活FX,而FX又是凝血级联反应的关键因素。来自Chugai的研究人员生成了一种IgG形状的bsAb,其几何形状与FVIII的结构和变构特性相似,从而恢复了凝血级联反应。这种分子被称为Hemliba®(Emizizumab),是两种经批准的bsAb之一。
细胞毒效应细胞靶向恶性组织的重定向是单克隆抗体的另一种作用方式。模拟抗体依赖性细胞介导的细胞毒性(ADCC)可能是消除癌细胞成熟的治疗策略之一。这个概念的基础是用治疗性抗体靶向癌细胞,然后通过Fc受体与NK细胞结合。NK细胞的激活反过来导致靶细胞死亡。Fc受体(FcγRIII,CD16)介导的作为bsAb功能的招募可以通过NK细胞表面的CD16结合到bsAb的Fc区域来实现,或者通过结合肿瘤特异性抗原和CD16的bsAb来实现。在这两种情况下,bsAb的组成和形式都会影响ADCC诱导的效果。用于Fc介导的NK细胞招募的bsAb的设计面临着这样一个挑战:所选择的形式需要确保Fc结构域对NK细胞CD16的有效结合。例如,单链抗体在N端或C端融合可显示出诱导ADCC的不同能力,从保留完全活性到ADCC活性的丧失。这些效应很可能是由于添加了结合区域和/或结合靶抗原对FcγRIII与NK细胞相互作用的空间阻碍引起的,不仅取决于形式,还取决于靶抗原(和表位)的选择,因此需要在个体基础上进行实验评估。通过与肿瘤细胞和CD16结合招募NK细胞的bsAb的ADCC效率同样依赖于结合抗体和形式的选择。例如,Reusch和他的同事已经证明,ADCC诱导的结合肿瘤相关抗原(TAA)CD30和CD16A的bsAb在双CD16A结合的TandAb(串联diabody)形式中比在diabody形式下的单价CD30/CD16A结合更优。
另一类特征明确且临床相关的用于效应细胞重定位的bsAb是T细胞结合的衍生物,它们是癌症免疫治疗的中心支柱。在这个领域,Blincyto®(blinatumomab)是代表,在II期研究中,这种CD19/CD3双特异性导致69%的复发/难治性B前体急性淋巴细胞白血病(ALL)患者完全缓解。
1985年首次证明T细胞双特异性(TCB)的概念后,出现了大量新的抗体形式。乍一看,两个细胞的交联似乎相当简单,不像前面提到的例子那样依赖于形式。然而,在测试各种抗体的过程中,很明显,设计安全有效的TCB不仅仅是一个即插即用的练习。例如,CD3结合抗体的高亲和力被证明与抗体迅速渗入次级淋巴组织和较低的肿瘤穿透率有关。其他人报告在注射高亲和力或二价CD3ε结合抗体后产生强烈的细胞因子释放效应。几何结构、价态和柔韧性也是影响T细胞招募治疗双抗的功能、效力和安全性的重要参数。
形式对T细胞接合双抗很重要。但为什么重要?要回答为什么某些抗体结构显示出比其他抗体更高的效力,就必须了解分子作用的生物学。
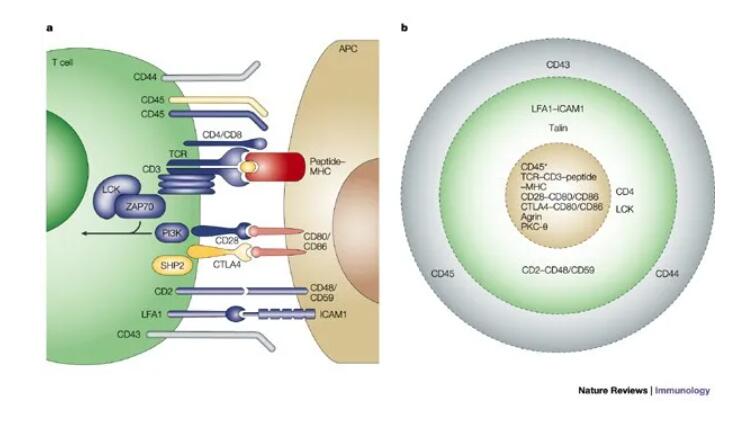
抗原呈递细胞(APC)与T细胞之间的生理免疫突触(IS)是T细胞活化的关键。在T细胞膜上,IS显示为一个嵌套的环状结构,常被称为“bullseye”。中心区cSMAC(中央超分子激活簇)包含诸如T细胞受体(TCR)复合物、共刺激分子CD28、抑制免疫检查点(如PD-1或CTLA-4)、信号介质如Lck(淋巴细胞特异性蛋白酪氨酸激酶)和PKC(蛋白激酶C),以及穿孔素等细胞毒性分子。cSMAC被周围的SMAC(pSMAC)包围,其中包含许多粘附分子,它们介导细胞-细胞的结合(如LFA1)。远端环(dSMAC)包括抑制性酪氨酸磷酸酶CD45和动态肌动蛋白。APC和T细胞之间的膜距离约为13–15nm。试验表明,一个拉长的膜间距降低了T细胞的活化,说明较近的距离是必须的,以适当地诱导T细胞活化。
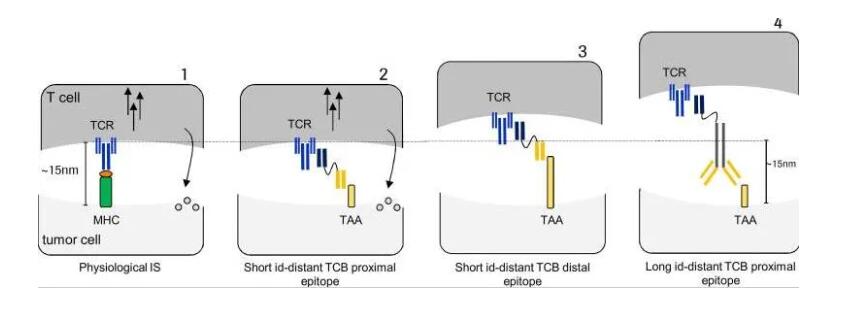
今天,众所周知,TCBs通过同时结合靶细胞上的TCR亚单位CD3ε和TAA来促进规则的细胞溶解免疫突触的形成。Offner等人用共聚焦显微镜证明,在生理性和抗体诱导的人工突触之间,来自所有SMAC环的突触特异性标记的共定位没有变化。Cartwright和他的同事利用定量荧光显微镜和纳米级右旋糖酐来分析哪些分子大小可以进入IS,以及是否存在引发IS排斥的大小阈值。他们确定可以证明4到13nm范围内的右旋糖酐可以穿透IS。另一方面,32-54nm之间的大分子被排除在外。他们的假设进一步被NK和肿瘤细胞之间免疫突触大小不同的抗体所证实。这些发现表明,人工免疫突触的距离很重要,在设计TCB时应该考虑到,TCB可能破坏而不是触发突触的形成。
Li和他的同事设计了一系列的TCB,这些TCB与FcRH5的不同区域结合,具有相似的结合亲和力。选择的表位位于靠近膜(近端)、远离膜(远端)或介于两者之间。骨髓瘤细胞与人T细胞和抗体共培养表明,靶向近端表位可诱导强的IS,并导致T细胞活化和肿瘤细胞溶解。考虑到这一机制,他们可以证明去除抑制性磷酸酶CD45会触发TCR磷酸化和T细胞信号的诱导。CD45是一种大的跨膜受体,其外结构域大小约为28-50nm。这反过来表明,较小的膜间距离有利于促进CD45从sSMAC中的清除。
Li等人分析了cSMAC内T细胞上的共抑制受体CTLA-4(细胞毒性T淋巴细胞相关蛋白-4)与其结合伴侣B7相互作用的结构基础。他们将细胞间的距离量化到14nm,这与其他人报道的免疫突触的间距一致。这一点对于产生拮抗抗体也很重要。例如,研究小组猜测,抗CTLA-4拮抗剂tremelimumab通过增加细胞间隙至15-19nm来破坏CTLA-4和B7之间的抑制轴,这两个受体就不能再相互作用了。
总的来说,人工免疫突触是一个高度组织化的结构,与生理拓扑结构具有相同的特征。肿瘤和T细胞之间的膜间距的发现表明,抗体尺寸是突触形成的一个关键参数。下图总结了开发人工IS(是否抗体模拟了生理性膜间距)或未有效形成(在远端表位结合或大抗体的情况下)的不同情况。
Wuellner等人比较了具有相同结合抗体和相同大小的两种双抗形式。种,FynomAb携带TAA结合抗体(即抗HER2 fynomer),N端融合到CD3的mAb上。在对照分子中,HER2结合抗体C端融合到Fc(图3A)。因此,TAA和CD3ε结合端之间的分子距离相差10-12nm。在肿瘤细胞和T细胞的共培养分析中,N端融合的CD3ε结合双抗诱导提高了20倍以上的效力(取决于靶细胞系,可以高达115倍),这表明TAA和TCR结合物之间的密切联系是有利的。这些观察结果与其他使用串联单链抗体靶向不同表位从而改变膜间距离的研究一致。
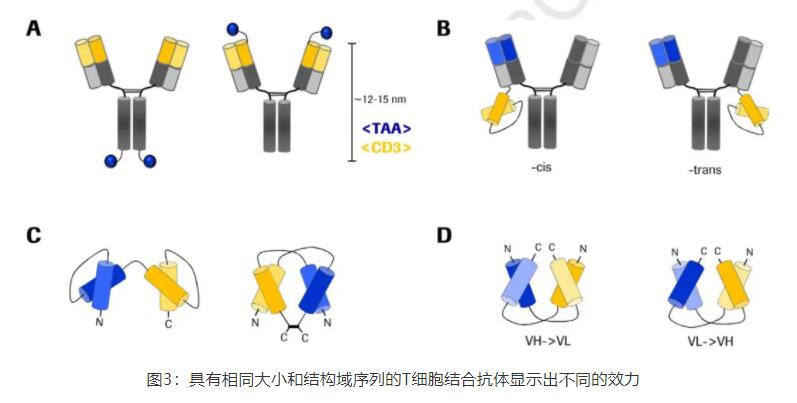
在一项类似的未发表的研究中,janssen对不同的bsAb形式进行了体外杀伤试验。在1+1格式中,CD3ε结合端位于一条重链上,而TAA结合端(centyrin)要么与Fc的C端融合,要么位于另一条重链上的相邻铰链区域。随着TAA与CD3ε结合端之间距离的增大,近距离结构的的效价比远距离高100倍。
近,Santich和他的同事解析了一个对称的双价bsAb,以探索价态和空间构型对bsAb诱导的T细胞毒性的重要性。他们使用单价肿瘤靶向IgG支架,将CD3ε靶向单链抗体以顺式(TAA结合端的轻链)或反式取向(非靶向结合端的轻链)融合到C-末端,得到1+1 bsAb(图3B)。他们可以证明顺式取向的分子在体外和体内都比具有更大的域间距离的反式分子更有效。顺式构型和短的结构域间距的有益增效其他人也有报道。重要的是,Santich等人强调进一步缩短结构域间距离通常是不利的,用2+2-BiTE-Fc分子进行的实验表明,CD3ε与肿瘤细胞之间的物理约束或机械耦合受损,导致T细胞反应降低。
Moore和他的同事比较了DART(双亲和重靶向)抗体与传统串联单链抗体(例如blinatumomab)在肿瘤细胞清除方面的效力。两种分子具有相同的分子量和相同的抗体序列。然而,这两种形式的区别在于分子的几何结构。在串联结构中,两个单链抗体一个接一个地位于一个链上,而DART分子在两个链(VLA-VHB,VLB-VHA)上以交替顺序携带两个特异性的可变域。此外,该结构在C-端的铰链区域中形成稳定的二硫键(图3C)。DART格式被证明在细胞毒性分析中的效力高出60倍。此外,DART分子在T细胞和靶细胞之间诱导了三倍多的细胞-细胞结合。活性的增加被解释为稳定的结构和支持双细胞结合的生理几何学。二硫键桥的另一个优点是增加了热稳定性。
Asano和他的同事发表了多个研究,集中在TCB背景下结构域定向对抗体功能的影响。他们表达了以EGFR和CD3ε为靶点的双特异性双抗体的所有四种可能的结构域顺序(图3D)。虽然所有的分子都被证明具有与两种TAA相似的结合能力,但肿瘤生长抑制作用却有显著差异。他们推测,由于其紧凑和不太灵活的结构,不同的diabody取向在细胞交联过程中面临不同的空间障碍,从而影响T细胞活性和随后的效应器功能。另一项研究通过重新排列基于Fc的diabodies中的结构域也进行了类似的观察,它们除了表现出不同的抗肿瘤作用外,还具有不同的抗降解性和体内半衰期。
Cheng等人研究了一种靶向黑色素瘤细胞的二唾液酸神经节苷脂(GD2)的TCB。他们观察到串联单链抗体形式的可变结构域方向(VH->VL或VL->VH)显著影响与抗原的结合力以及肿瘤和T细胞存在时的细胞毒性效应。根据分子模型和抗原对接数据,证实了CDR环的关键残基在VH和VL方向上具有不同的构象,这些残基对抗原结合起着重要作用。
TriFab Contorsbody:紧凑的架构具有更大的潜力
近发表了一种抗体形式,名为TriFab,其中Fc部分的CH2结构域被T细胞结合域取代。如图4A所示,CD3ε结合端的可变区被并入抗体的主干区,而不是像在大多数TCB形式中发现的N-或C-末端融合。
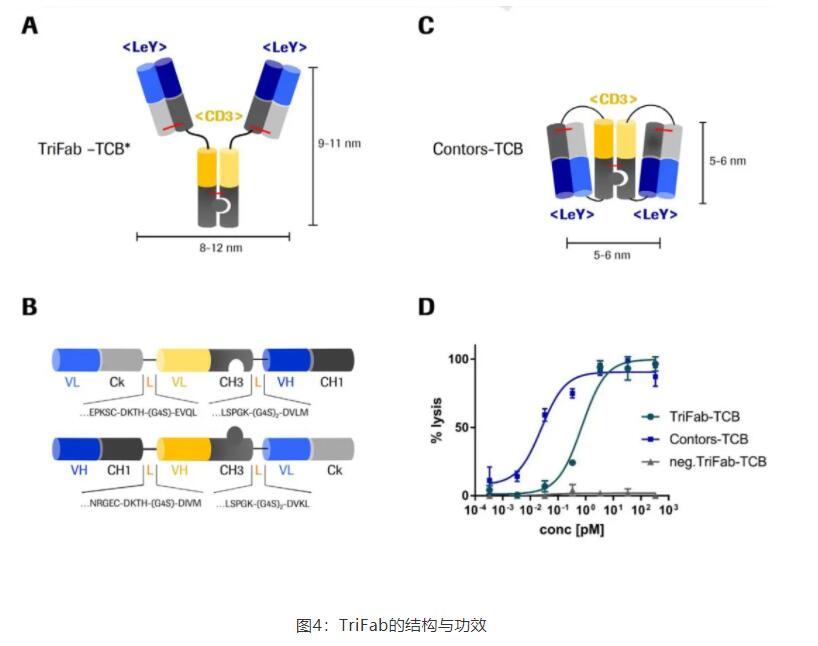
另一种与TriFab和Contorsbody架构相关的新颖格式是“TriFab Contorsbody”。这个分子是由两个链(图4B)组成的三价分子(图4C)。CD3ε结合基团呈反式排列。在与人PBMC共培养试验中比较两种TCB形式的疗效,发现Contors TCB比TriFab形式活性高35倍(图4D)。
在Contorsbody中,两个靶向部分(蓝色)更接近,这可能导致靶细胞膜上更高的局部聚集。因此,与规则的Y形IgG样分子相比,TCR的同时反式结合将导致TCR更密集的聚集。这种活性的增加可以用permissive几何模型解释,该模型描述了由于细胞质CD3ε结构域的诱导结构改变,TCR聚集导致活化增强。
与IgG形状的分子相比,Contorsbody结构更紧凑。Y形结构的回转半径约为6nm,而Contorsbody的回转半径约为3nm;将半径除以2等于体积减小了8倍。正因为如此,Contorsbody可能更适合穿透IS。如Cartwright等人所述,这会导致更高的局部浓度。从而导致细胞毒性增强。
TriFab-Contorsbody格式不仅使两个结合基团更加紧密地结合在一起,而且在理想的trans结构中提供了第三个结合基团。使CD3ε结合更容易,这使得T细胞和靶细胞可以同时有效结合。
到目前为止,两种双特异性抗体已经上市,然而,大量正在进行的临床试验可能预示着还有更多的双特异性抗体。除此之外,两种不同抗原的同时结合在许多不同的治疗领域是相关的,不仅限于从两个现有的单一特异性抗体的联合治疗演变而来共同阻断策略。设计和选择合适的结合抗体形式组合对于生成具有所需功能的bsAb至关重要。实现这一目标没有“标准程序”,而是必须根据潜在的生物学和结构条件逐个解决。
Blinatumomab的成功诱发了T细胞双特异性抗体开发的热潮。许多与T细胞有关的抗体以各种形式和几何结构得到测试,很明显,它们的效力和安全性各不相同。同时,通过显微镜和结晶学的研究,揭示了免疫突触形成的结构基础。结合这些知识和许多临床试验的结果,现在可以解释为什么有些形式比其他形式更有效。这些经验教训将有助于设计安全有效的T细胞活化的双特异性抗体。相信,未来基于结构优化设计的双特异性抗体会越来越多地出现在临床研究中。
参考文献:
Format and geometries matter: Structure-based design defines the functionality of bispecific antibodies. Comput Struct Biotechnol J. 2020 May 14;18:1221-1227.



